The role of NEP1-like proteins in plant-pathogen interaction
Necrosis- and ethylene inducing protein 1-like proteins (NLPs) are produced by phytopathogenic bacteria, fungi and oomycetes. Many of them are cytolytic toxins that promote virulence of the pathogens with necrotrophic life style, making nutrients available by disruption of host tissue (1). One prominent NLP-producing organism is Phytophthora infestans, causing agent of the great famine in Ireland in the middle of the 19th century (Fig. 1).

Fig. 1: Diseases caused by NLP-producing pathogens:
Late blight (A) by Phytophthora infestans and tuber rot (B) by Pectobacterium carotovorum on potato.
NLPs facilitate cell lysis in dicotyledonous plants, macroscopically visible as necrosis. NLPs are structurally related to pore-forming toxins from marine invertebrates, acting on sphingomyelin-containing animal membranes. Likewise, NLPs bind to plant-specific sphingolipids, glycosyl-inositol-phosphoryl-ceramides (GIPCs) (2). GIPCs occur in high numbers in the outer leaflet of the plant plasma membrane and have a variable glycosylation pattern amongst plant species. Whereas NLP-sensitive dicots have mainly GIPCs with two terminal hexoses (Fig. 2 A), many monocotyledonous plants have a majority of longer, more complex glycosylated GIPCs (Fig. 2 B). The length of this glycosylation pattern determines the distance of the bound NLP to the membrane surface, thus the accessibility of the membrane for the toxin insertion (Fig. 2 C) and subsequent cytolysis (Fig. 2 D).
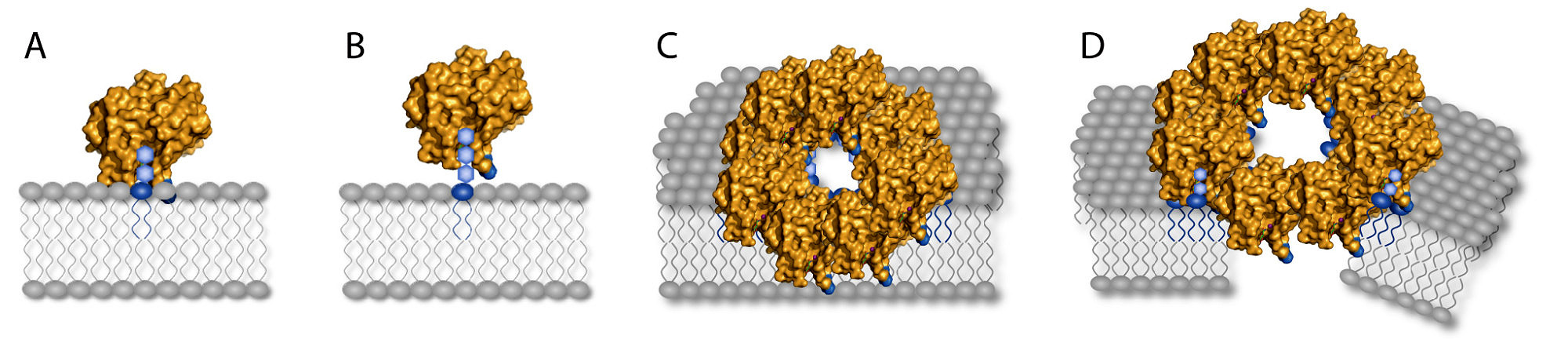
Fig. 2: Model of NLP-membrane interaction:
NLPs bind to glycosylated lipids in outer membrane leaflet (A: NLP-sensitive, B: NLP-insensitive). In case of close contact to the membrane (A), Oligomerization leads to perforation (C) and disruption of the plasma membrane (D).
Despite the deleterious action of NLPs, some plants evolved mechanisms to detect NLPs and induce defence and counteract infection at NLP concentrations that precede necrotic concentrations. While the cytolytic activity itself induces plant immunity, some Brassicaceae additionally detect a short peptide within the NLP protein via a cell surface receptor protein, RLP23 (3; 4).
Our research focuses on determinants of plants being sensitive or insensitive to NLPs. Understanding the mechanism of NLP-triggered membrane disruption enables new strategies for the control of plant diseases and the engineering of NLPs as bioherbicides.
Ottmann, C., Luberacki, B., Küfner, I., Koch, W., Brunner, F., Weyand, M., Mattinen, L., Pirhonen, M., Anderluh, G., Seitz, H.U., Nürnberger, T., and Oecking, C. (2009). A common toxin fold mediates microbial attack and plant defense. Proc Natl Acad Sci U S A 106, 10359-10364.
Lenarcic, T., Albert, I., Böhm, H., Hodnik, V., Pirc, K., Zavec, A.B., Podobnik, M., Pahovnik, D., Zagar, E., Pruitt, R., Greimel, P., Yamaji-Hasegawa, A., Kobayashi, T., Zienkiewicz, A., Gomann, J., Mortimer, J.C., Fang, L., Mamode-Cassim, A., Deleu, M., Lins, L., Oecking, C., Feussner, I., Mongrand, S., Anderluh, G., and Nürnberger, T. (2017). Eudicot plant-specific sphingolipids determine host selectivity of microbial NLP cytolysins. Science 358, 1431-1434.
Albert, I., Böhm, H., Albert, M., Feiler, C.E., Imkampe, J., Wallmeroth, N., Brancato, C., Raaymakers, T.M., Oome, S., Zhang, H.Q., Krol, E., Grefen, C., Gust, A.A., Chai, J.J., Hedrich, R., Van den Ackerveken, G., and Nürnberger, T. (2015). An RLP23-SOBIR1-BAK1 complex mediates NLP-triggered immunity. Nat Plants 1.
Böhm, H., Albert, I., Oome, S., Raaymakers, T.M., Van den Ackerveken, G., and Nürnberger, T. (2014). A Conserved Peptide Pattern from a Widespread Microbial Virulence Factor Triggers Pattern-Induced Immunity in Arabidopsis. Plos Pathog 10.